
Organografia vegetal/Flor
|
|
Aquesta pàgina s'està elaborant i està inacabada.
L'usuari Simonjoan hi està treballant i és possible que hi trobeu algun defecte. Us preguem paciència, però si voleu editar, comenteu abans qualsevol canvi. Aquest avís és temporal, utilitzeu-lo només mentre esteu treballant activament en un article, com a molt durant uns dies. |
Una flor és l'estructura reproductiva característica de les plantes espermatofitines o fanerògame. La funció de la flor és produir llavors a través de la reproducció sexual. Per les plantes, les llavors són la pròxima generació, i serveixen com el principal mitjà a través del qual es perpetuen i propaguen les espècies.
Tots els espermatofitins tenen flors que produeixen llavors, però l'organització interna de la flor és molt diferent en els dos principals grups d'espermatofitins: les gimnospermes vivents i les angiospermes. Les gimnospermes poden tenir flors que es reuneixen en estròbils, o la mateixa flor pot ser un estròbil de fulles fèrtils. En canvi, una flor típica d'angiosperma està composta de quatre tipus de fulles modificades, tant estructuralment com fisiològica, per produir i protegir els gàmetes: sèpals, pètals, estams i carpels.[1] Després de la fertilització, la flor dóna origen, per transformació d'algunes de les seves parts, a un fruit que conté les llavors.[2] El grup de les angiospermes, amb més de 250.000 espècies, és un llinatge evolutivament reeixit que conforma la major part de la flora terrestre existent. La flor és el caràcter definidor del grup i és probablement un factor clau en el seu èxit evolutiu. És una estructura complexa el pla organitzacional de la qual és conservat en gairebé tots els membres del grup, tot i que presenta una enorme diversitat en la morfologia i fisiologia de totes i cadascuna de les parts que la componen. La base genètica i adaptativa d'aquesta diversitat s'està començant a comprendre en detall,[3] igual que el seu origen, que data del Cretaci inferior, i la seva posterior evolució en estreta interrelació amb els animals que s'encarreguen de transportar els gàmetes.
Amb independència dels aspectes ja mencionats, la flor és un objecte important pels éssers humans. A través de la història i de les diferents cultures, la flor sempre ha tingut un lloc a les societats humanes, sigui per la seva bellesa intrínseca o el seu simbolisme. De fet, es cultiven espècies per obtenir-ne flors des de fa més de 5.000 anys i, avui en dia, aquest art s'ha transformat en una indústria en contínua expansió: la floricultura.
Morfologia
[modifica]

Una flor és un brot o branca de creixement limitat que porta al seu àpex un nombre variable de fulles modificades anomenades antofil·les. L'eix d'aquest brot és generalment molt curt, amb entrenusos molt propers, de manera que sembla que les antofil·les s'insereixen al mateix nivell. la flor que no presenta les peces homòlogues al mateix nivell de l'eix floral, sinó disposades en una línia helicoïdal s'anomena acíclica, i cíclica la que presenta les peces homòlogues als mateixos nivells de l'eix, essent cada nivell un verticil.
A la flor s'hi pot trobar dos tipus bàsics d'antofil·les:
- Estèrils, que formen el periant. Generalment el periant està format per dos tipus de peces:
Quan entre les fulles estèrils no es pot diferenciar calze i corol·la, s'utilitza el terme perigoni (en comptes de periant) i a les seves peces se les anomena tèpals (per contraposició a sèpals i pètals). Hi ha flors sense peces estèrils, i llavors se'n diuen flors aclamídies.
- Fèrtils, que són els elements essencials de la flor ja que són les fulles portadores d'esporangis i on té lloc la funció reproductora de tota la planta. Poden ser de dos tipus:
Les flors s'originen a partir de brots que es troben l'axil·la de les fulles (molt sovint transformades en bràctees). El més habitual és que les flors es presentin agrupades en conjunts més o menys aparents anomenats inflorescències tot i que també les podem trobar separades com a flors solitàries. De vegades és difícil distingir una inflorescència d'una flor. És el cas de la inflorescència del dent de lleó (Taraxacum officinale), que sembla una flor solitària quan, en realitat, es tracta d'una inflorescència en capítol.
Eix o tàlem floral
[modifica]Totes aquestes peces es disposen sobre l'eix floral que de vegades s'insereix directament a la tija (flor sèssil) i d'altres ho fa mitjançant un pedicel o peduncle més o menys llarg (flor pedunculada). Sovint a la base del pedicel hi ha la fulleta transformada en una bràctea floral. La disposició de les peces pot ser:
- Acíclica: les peces es troben l'una a continuació de l'altre, amb entrenusos molt curts, i de manera helicoïdal. Aquest tipus de flor és una flor helicoïdal o acíclica, com la de la magnòlia.
- Cíclica, en verticil, de manera que totes les peces d'un mateix tipus surten d'un sol nus. En aquest cas hi ha una flor verticil·lada o cíclica, que poden tenir més d'un verticil de cada tipus de peces florals. L'esquema general, però, consta de quatre verticils, els dos primers corresponents a les peces del periant (calze i corol·la) i el tercer i quart a l'androceu i el gineceu, respectivament.
L'eix floral és allargat i més o menys cònic a les gimnospermes i les angiospermes primitives (com Magnolia), però en el decurs de l'evolució s'ha anat escurçant fins a esdevenir pla o còncau i, en aquest cas, s'anomena receptacle floral. Aquests receptacles còncaus solen soldar-se amb la base del gineceu.

|

|
Simetria
[modifica]Si es deixen de banda les flors helicoïdals, la major part de les flors cícliques tenen les peces, especialment les del periant, disposades de manera simètrica. Segons la simetria, les flors es classifiquen en:
- Flors actinomorfes (també dites regulars o radiades), amb més de dos plans de simetria. Hi ha molts exemples a la nostra flora, com ara Borago officinalis, Hepatica nobilis, Caltha palustris (a la imatge),...
- Flors dissimètriques amb dos plans de simetria perpendiculars i no equivalents. Un exemple és Hypecoum imberbe [1], però no són gaire freqüents, tot i que la flor de les brassicàcies ho seria si prenem en consideració l'androceu a més del periant.
- Flors zigomorfes, amb un únic pla de simetria, com ara la flor de les lamiàcies i les fabàcies.
- Flors asimètriques, si no tenen cap pla de simetria com, per exemple, Canna indica (la canya d'Índies, utilitzada en jardineria).
-
 Flor actinomorfa
Flor actinomorfa
(Caltha palustris) -
 Flor zigomorfa
Flor zigomorfa
(Lotus corniculatus) -
 Flor asimètrica
Flor asimètrica
(Canna indica)



Repartició de sexes
[modifica]La major part de les flors són hermafrodites, és a dir, tenen gineceu i androceu alhora. Això no obstant, també són freqüents les flors unisexuals, que només tenen gineceu (flors femenines) o només androceu (flors masculines). Entre les gimnospermes les flors solen ser unisexuals; en canvi, les flors de les angiospermes són, habitualment, hermafrodites, exceptuant alguns grups en els quals la pol·linització és anemòfila.
Segons la distribució de flors a la planta hi ha:
- Plantes monoiques, amb flors hermafrodites o els dos tipus de flors unisexuals al mateix peu. Exemple: Quercus ilex (alzina), Pinus (pins),...
- Plantes dioiques, amb flors unisexuals a peus separats. Exemple: Salix (salzes).
- Plantes polígames, amb flors hermafrodites i unisexuals al mateix peu. Exemple: Fraxinus (freixe) o Ceratonia siliqua (garrofer).
-
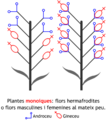 Plantes monoiques
Plantes monoiques -
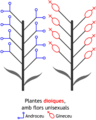 Plantes dioiques
Plantes dioiques -
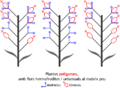 Plantes polígames
Plantes polígames



Representació floral
[modifica]
Exemple: * K4 C4 A4+2 G(2)
Diplotaxis erucoides, flor amb:
- Simetria central (actinomorfa)
- Calze amb quatre sèpals lliures
- Corol·la amb quatre pètals lliures
- Sis estams tetradínams
- Gineceu bicarpel·lar súper amb dos carpels soldats
Per representar les característiques fonamentals d'una flor es pot utilitzar:
- La fórmula floral, un conjunt de símbols, lletres i números que expliquen la simetria de la flor, el nombre de peces per verticil, la soldadura de les peces, la posició de l'ovari,... Per a més informació consultar [2]
- El diagrama floral, una projecció de la flor on les peces més externes corresponen als verticils inferiors. S'hi representen el nombre de peces de cada mena, la soldadura i la posició relativa.
-
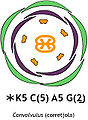 Diagrama i fórmula floral de Convolvulus
Diagrama i fórmula floral de Convolvulus -
 Convolvulus arvensis.
Convolvulus arvensis.


Evolució de la flor
[modifica]Les primeres llavors fòssils (produïdes dins flors) provenen del Devonià, període en el qual les plantes dominants eren els pteridòfits. Els espermatòfits o plantes amb flors provenen d'un grup de pteridòfits que va desaparèixer cap a finals del Devonià.
Les flors es van originar per segregació de les fulles fèrtils, portadores d'esporangis, que es van anar agrupant helicoïdalment a l'extrem de branques curtes. Progressivament aquestes branques i els entrenusos es van anar escurçant i van aparèixer altres modificacions, com ara un periant que embolcallava les fulles fèrtils. La taula següent recull les característiques florals considerades "primitives" envers les considerades "avançades".
| Primitives | Avançades |
|---|---|
|
|
| Segons HEYWOOD, Vernon H. Las plantas con flores. Barcelona: Editorial Reverté, 1985. 332 p. ISBN: 84-29119205. | |
Desenvolupament floral
[modifica]Un moment crític en el cicle vital de les plantes amb flor és aquell en què realitzen la transició de la fase juvenil a la fase reproductiva. Per aconseguir reproduir-se amb èxit, és imprescindible, d'una banda, que la floració es produeixi en sincronia amb altres membres de la mateixa espècie i, d'altra banda, que la planta hagi assolit un cert nivell de creixement que li permeti afrontar la despesa energètica implicada en el procés de formació de flors, fruits i llavors. Per tant, la planta ha d'integrar informació ambiental i endògena abans de patir la transició. Entre els factors externs que més influeixen en la floració es troben la llum (tant la intensitat lumínica com la duració del lapse diari d'hores de llum o fotoperíode) i la temperatura. Aquest últim factor té un paper doble. D'ua banda, les temperatures més altes incrementen la taxa de creixement i diferenciació de les plantes, accelerant-ne la transició al període de floració. D'altra banda, moltes plantes necessiten ser exposades transitòriament a temperatures molt baixes durant la fase vegetativa o juvenil per poder florir, un procés anomenat vernalització. A més d'aquests factors externs, també tenen un paper important factors endògens com el rellotge circadià i algunes hormones com les giberel·lines, l'àcid salicílic i l'àcid abscíssic. Així doncs, la planta model Arabidopsis thaliana floreix més ràpidament quan els dies s'allarguen (es diu que és una planta de dia llarg), ja ha patit la vernalització (ha estat exposada al fred durant l'hivern) i la temperatura ambient augmenta. És a dir, igual que una miríada d'altres espècies, la floració d'Arabidopsis és induïda per les condicions que s'observen a la primavera.[4]
L'anàlisi genètica de mutants amb el temps de floració alterat ha permès diseccionar la inducció de la floració en almenys quatre vies de senyalització: la primera s'activa quan els dies són llargs; la segona funciona fonamentalment quan els dies són curts i depèn de les giberel·lines; la tercera participa en la regulació de la floració en resposta a la temperatura ambiental; i la quarta està implicada en la vernalització.[5]
Quan la planta arriba a l'estat reproductiu del desenvolupament, algun o tots els meristemes apicals del seu brost deixen d'iniciar fulles normals i comencen a produir parts florals segons la seqüència característica de l'espècie en qüestió i amb un nombre variable de bràctees situades entre les fulles i la flor. En aquest procés, els meristemes apicals passen de tenir un creixement indeterminat a tenir-ne un de determinat, car la formació de la flor és generalment l'esdeveniment final de l'activitat d'un meristema apical. En les plantes anuals, l'advent de l'estat reproductiu també significa un acostament a la consumació del cicle de vida en la seva totalitat. En les plantes perennes, la floració es repeteix un nombre variable de vegades que depèn de la longevitat de l'espècie.
Una flor es pot originar a l'àpex del brost principal o al de les branques laterals, o als dos llocs alhora. En moltes espècies, el canvi cap a la floració inclou la formació d'una inflorescència. Un fenomen observat sovint durant la iniciació de l'estat reproductiu és un brusc i ràpida allargament de l'eix. Aquest creixement és particularment important en plantes d'hàbit en roseta durant el període vegetatiu, com ara les gramínees o les plantes bulboses.[6]
Els òrgans florals s'inicien com a fulles normals per divisions periclinals de cèl·lules situades més o menys profundament per sota de la protodermis o a la mateixa protodermis.[7][8][9] Les divisions inicials periclinals són seguides per altres, incloent-hi d'anticlinals, i el primordi es converteix en una protuberància. Els sèpals s'assemblen més a les fulles normals al seu inici i desenvolupament. El tipus de creixement dels pètals també és més o menys similar al de les fulles. Els estams s'originen com a estructures gruixudes i curtes, desenvolupant-se després el filament per creixement intercalar. El desenvolupament del gineceu varia en detall en relació amb la unió dels carpels entre si i amb altres parts florals. Si els carpels no estan units, el primordi del carpel individual s'inicia en un punt per divisions periclinals a l'àpex, que després es propaguen adaxialment. El creixement cap amunt, comparable al marginal en una fulla, eleva el carpel com una estructura en forma de bossa. Als ovaris sincàrpics els carpels s'originen com a primordis individuals o conjuntament com una unitat estructural.[6]
Biologia molecular del desenvolupament floral
[modifica]
La definició de flor mencionada al principi d'aquest article s'emmarca dins del context de l'anomenada "teoria foliar", que suggereix que totes les peces florals són fulles estructuralment modificades i funcionalment especialitzades en la reproducció o la protecció. La formulació d'aquesta teoria començà el 1790 quan Goethe publicà la seva obra Un intent d'interpretar la metamorfosi de les plantes (Versuch die Metamorphose der Pflanzen zu erklaren).[10] En aquest llibre, que consistia en 123 paràgrafs enumerats, Goethe escrigué:
Gairebé 200 anys després d'aquesta publicació, la biologia molecular i la biologia evolutiva del desenvolupament han inventat eines per descodificar l'estructura molecular bàsica del desenvolupament floral. Una de les fites d'aquestes disciplines ha estat la descripció de l'anomenat model "ABC", un model genètic simple i elegant per explicar la determinació de la identitat dels òrgans florals,[11] confirmat pel descobriment de la profunda homologia i conservació dels gens implicats en el model, pertanyents a tàxons que divergiren ja fa milions d'anys.[12][13]
El model ABC del desenvolupament floral fou proposat com a model explicatiu del conjunt de mecanismes genètics que donen lloc a l'establiment d'identitats d'òrgan en Arabidopsis thaliana, com a representant de les ròsides i d'Antirrhinum majus, pertanyent al clade de les astèrides. Ambdues espècies tenen quatre verticils (sèpals, pètals, estams i carpels), definits per l'expressió diferencial a cada verticil d'una sèrie de gens homeòtics. El model ABS estableix que la identitat dels òrgans presents en aquests quatre verticils florals és conseqüència de la interacció de com a mínim tres tipus de productes gènics de funcions diferents. D'aquesta manera, els sèpals es caracteritzen per l'expressió exclusiva de gens A; als pètals, en canvi, es coexpressen els gens A i B; als estams, els gens de funció B i C estableixen la seva identitat; i als carpels només cal l'activitat de gens de funció C. A més, els gens A i C són antagonistes recíprocs.[14]
El fet que aquests gens homeòtics determinen la identitat d'òrgan es posa de manifest quan un gen representant d'una determinada funció, per exemple l'A, no s'expressa. Així, en Arabidopsis aquesta pèrdua causa l'aparició d'una flor composta per un verticil de carpels, un altre d'estams, un segon d'estams, i un segon de carpels.[14] S'ha proposat, a més de les ja mencionades funcions A, B i C, l'existència d'unes de suplementàries: la D i l'E. La funció D especificariala identitat de l'òvul, com una funció reproductiva separada del desenvolupament dels carpels i posterior en aparició a la determinació d'aquests últims.[15] La funció E correspondria a un requisit fisiològic propi de tots els verticils florals, tot i que inicialment fou descrita com a necessària pel desenvolupament dels tres verticiles més interns (funció E sensu stricto).[16] No obstant això, la seva accepció sensu lato suggereix que és requerit als quatre verticils.[17] D'aquesta manera, quan es perd la funció D, els òvuls passen a ser estructures similars a fulles, i quan es perd la funció E sensu stricto, els òrgans florals dels tres verticils més interns es transformen en sèpals,[16] mentre que, si es perd la funció E sensu lato, tots els verticils són similars a fulles.[17][18]
Així doncs, el model ABC explica els aspectes més importants del desenvolupament floral, des dels més teòrics (una posada al dia de les idees de Goethe) fins als més pràctics, com ara la manipulació dels gens ABC per produir flors amb qualsevol tipus de combinació de peces florals.[19][10]
Les flors comestibles
[modifica]Les flors no són una part important de la nostra dieta, ho són més els fruits, les llavors o les fulles. Això no obstant sí que en consumim les flors o parts de les flors d'algunes plantes. La carxofa, per exemple, és la inflorescència de Cynara scolymus; la coliflor i el bròquil són també inflorescències de Brassica oleracea; les tàperes són poncelles florals de Capparis spinosa i el safrà els estigmes de Crocus sativus. També es mengen a algunes contrades les flors masculines de carbassó (Cucurbita pepo).
D'altres flors es prenen en infusió, com ara la camamilla (Matricaria chamomilla) o la til·la (Tilia platyphyllos).
Més rarament en fem melmelades o caramels, com els de violeta (Viola alba).
Referències bibliogràfiques
[modifica]- FONT I QUER, Pius. Iniciació a la Botànica. 2a edició a cura d'Oriol de Bolòs. Barcelona: Editorial Fontalba, 1979. 262 p. ISBN: 84-85530-08-X.
- HEYWOOD, Vernon H. Las plantas con flores. Barcelona: Editorial Reverté, 1985. 332 p. ISBN: 84-29119205.
- Història Natural dels Països Catalans. Volum 6: Plantes superiors. Barcelona: Fundació Enciclopèdia Catalana, 1988. 463 p. ISBN: 84-7739-015-0.
Referències
[modifica]<referències/>
Enllaços externs
[modifica]- Botànica agrícola i forestal Web de botànica elaborada per la Universitat de Lleida i la Universitat Politècnica de Catalunya.
- ↑ Universitat d'Hamburg. Departament de Biologia. Botànica en línia.Flower Morphology of Plants. Consultat el 5 d'abril del 2009;
- ↑ Plantilla:Cite book
- ↑ Damerval, C.; Nadot, S. 2007. Evolution of Perianth and Stamen Characteristics with Respect to Floral Symmetry in Ranunculales. Ann. Bot. 100: 631-640.
- ↑ Plantilla:Cite web
- ↑ Simpson G. G., Gendall A. R. & Dean C. 1999. When to switch to flowering. Annu. Rev. Cell Dev. Biol. 15: 519-50.
- ↑ 6,0 6,1 Esau, K. 1988. Anatomía de las plantas con semilla. Ed. Hemisferio Sur. ISBN 950-004-233-9
- ↑ Tepfer, S. 1953. Floral anatomy and ontogeny in Aquilegia formosa and Ranunculus repens. Univ. Calif. Pubs. Bot. 25:513-648.
- ↑ Tucker, S.C. 1959. Ontoeny of the inflorescence and the flower of Drymis winteri. Univ. Calif. Pubs. Bot. 30:257-336.
- ↑ Barnard, C. 1957. Floral histogenesis in the monocotyledons. I. The Gramineae, Aust. J. Bot. 5:115-128.
- ↑ 10,0 10,1 Dornelas, M. C.; Dornelas, O. 2005. Da folha à flor: revisitando os conceitos de Goethe sobre a ¨metamorfose¨ das plantas. Braz. J. Plant Physiol., Londrina, v. 17, n. 4,
- ↑ Coen E. S., Meyerowitz E. M. (1991) The war of the whorls: genetic interactions controlling flower development. Nature 353:31-37.
- ↑ Theissen G, Becker A, Di Rosa A, Kanno A, Kim JT, Munster T, Winter KU, Saedler H. (2000) A short history of MADS-box genes in plants. Plant Mol. Biol. 42:115-149.
- ↑ Irish VF, Litt A (2005) Flower development and evolution: gene duplication, diversification and redeployment. Curr. Opin. Gen. Dev. 15:1-7.
- ↑ 14,0 14,1 Bowman, J. L. et al.. Expression of the Arabidopsis floral homeotic gene AGAMOUS is restricted to specific cell types late in flower development. Plant Cell 3, 749-758
- ↑ L. Colombo, J. Franken, E. Koetje, J. van Went, H. J. Dons, G. C. Angenent i A. J. van Tunen. The petunia MADS box gene FBP11 determines ovule identity. Plant Cell. Novembre del 1995; 7(11): 1859–1868.
- ↑ 16,0 16,1 Soraya Pelaz, Gary S. Ditta, Elvira Baumann, Ellen Wisman and Martin F. Yanofsky. B and C floral organ identity functions require SEPALLATA MADS-box genes Nature 405, 200-203 (11 de maig del 2000) | doi:10.1038/35012103
- ↑ 17,0 17,1 Plantilla:Cite journal
- ↑ Weigel D., Meyerowitz E. M. (1994) The ABCs of floral homeotic genes. Cell 78:203-209.
- ↑ Dornelas M. C. (2000) Construindo flores: O controle molecular da arquitetura floral. Biotecnol. Ciênc. Desenvolv. 12:44-46.